Science
General
Science is an activity by which man seeks to understand the universe, i.e. all the observed phenomena. Scientists do this by making generalizations (or laws) about the phenomena and providing explanations (or theories) about the generalizations. That science seeks laws and theories implies two assumptions about the universe: (a) it is consistent in time and space—otherwise generalizations would be impossible and (b) it is understandable—otherwise theories would be impossible. These two assumptions are themselves testable hypotheses. A third assumption that it is important to state is (c) the observed phenomena accurately reflect the underlying reality of the universe (the true nature of things.)
The Logical Basis Of Scientific Testing
The key to the success of science is that it subjects its ideas to empirical tests. The basis of the testing process is diagrammed in figure 1. Observed phenomena are what we detect; they are produced by the underlying true nature of things which we seek to discover. Because these "true natures" are not directly observable, our hypotheses about them cannot be tested by direct observation. Thus one deduces observable logical consequences (the predicted phenomena) which may be compared with observations.
If the consequences do not match the observations, then the hypothesis does not match the true nature of things and we say that the hypothesis has been falsified. If they do match, we cannot be sure that the hypothesis matches the true nature of things, for experience has shown us that several hypotheses may equally well explain a set of phenomena and we may have to look further to distinguish them. Also further investigations may reveal new phenomena that falsify a previously accepted hypothesis. Thus an hypothesis may be disproved or falsified, but never proved.
The temporal sequence of this activity is illustrated in figure 2. There is a problem to be solved. One guesses at the answer—the hypothesis—and deduces the observable consequences which are then compared with new observations. (It is most desirable that the hypothesis have consequences for an array of phenomena that were not considered in its original formulation. This independence of testing from the original basis is important to avoid confirming the hypothesis by circular arguments.) The hypothesis may be rejected, accepted or modified. As we go around this cycle of intellectual activity, we hopefully come closer and closer to the truth—defined by the philosopher Karl Popper as correspondence of the consequences with the observations.
The Distinction Between Laws And Theories
A law is the assertion of an invariable association (not necessarily sequential or causal) of observations. It can be tested by direct measurements on the relevant phenomena. A theory is an explanation of laws (i.e. expresses them in a more acceptable and satisfactory form—more satisfactory because the form is more familiar and/or general.) Almost always, some of the ideas in a theory cannot be tested by direct perception (an important distinction from laws!) A theory may postulate entities or processes too small to be observed (atoms, electrons, etc.), intrinsically invisible (electromagnetic fields, curved space, black holes, etc.), too far distant (stars, etc.), inaccessible to observations (the interior of the earth, sun, etc.) or remote in time (evolution.)
The Properties Of Scientific Hypotheses
From the above discussion we see that scientific hypotheses must be testable (falsifiable). Because the goal of science is understanding, they should also be informative, providing the how, where, when and why a phenomenon occurs and what it is or what causes it. The more informative a hypothesis, the more specific the logical consequences and the greater is the potential for falsifiability. It is also preferred that a given hypothesis cover as wide a range of phenomena as possible (Whewell's consilience) with the fewest assumptions or the simplest possible mechanism (i.e. the principle of Occam's razor). Generating the most logical consequences is important, because the testing of these constitutes a guide for future research, pointing out what facets of nature should be investigated next. This role is so important that Popper advises that an apparently falsified hypothesis should not be discarded until all of its logical consequences have been explored. This insures that we will have the maximum knowledge necessary upon which to base a new hypothesis. Thus fruitful hypotheses can make important contributions to scientific knowledge even if they eventually turn out to be false! Whewell considered consilience the definitive evidence for the truth of a theory.
By now it should be clear that vitalistic and supernatural hypotheses that invariably postulate vague and amorphous mechanisms whose workings are beyond human comprehension are untestable and uninformative and hence not scientific. In fact they are not even explanations, but statements of unsolvable mysteries beyond the powers of scientific investigation.
The Need For Specific Conditions
Actual testing is more complex than we have described (see figure 3) because any general hypothesis (law or theory) can only entail general consequences. But it can only be tested against specific instances of real systems. Thus we need a set of initial or specific conditions of the system under test. Combined with the general postulates, they constitute a model—an abstract representation of some natural or artificial system whose predicted behavior is compared with the real system.
Several important points can be made here:
In carrying out such a test, we are testing several things at the same time: (1) the general law or theory, (2) the correctness of the specific conditions, (3) the deductive processes leading to the consequences and (4) the accuracy of the observations. Difficulties with any of these can lead to apparent falsification. Thus it is said that when Newton was first testing his gravitational hypothesis using the specific instance of the moon's orbit in 1665, the calculations did not agree with the observations because, as it turned out, he was using an incorrect value for the moon's distance. In recent attempts to improve upon the orbital parameters of the planet Neptune, possible inaccuracies in early observations of the planet and in the specific conditions, including the possible perturbations of a tenth planet (other than Pluto) had to be considered. In piecing together the evolutionary history of a particular organism, say man, each new fossil adds new observational details to be considered by the specific conditions. Here the specific conditions constitute what Philip Kitcher calls a "Darwinian history" or "evolutionary scenario" accounting for the known facts about fossil and recent men.
We can't test every relevant instance—only those for which we have sufficient knowledge of the specific conditions. If, for example, we have only one observation on a newly discovered asteroid, we cannot compute its specific elliptical orbit and thus its future positions in order to test Kepler's and Newton's laws. Likewise, without specific fossil evidence, we can only speculate on the evolutionary pathways leading to such oddities as the archerfish and the platypus.
Predictions
The observable logical consequences are often called predictions and creationists often point out that evolutionists cannot predict the evolutionary future, thus evolution is not testable. But these consequences are necessarily predictions only in the broad sense that they predict the results of observations yet to be made. They are not necessarily predictions of the future state of some natural system. In fact, most scientific "predictions" are conditional: given the relevant initial conditions, usually of some artificial system (such as a mixture of chemicals in a test tube) the immediate future state can be predicted. It is different with a natural system which is usually very complex and where most of the specific conditions are not under our control or are unknown. Only in the very special cases where the system is fairly isolated and repetitive (such as the movements of the bodies of the solar system and phenomena related to them like the seasonal climatic patterns; or the life cycles of organisms) can we normally make successful predictions of the future. (Consider the success of the weatherman in predicting the immediate future weather!) Thus the inability of Darwinism to predict the future evolution of species does not set it apart from other scientific hypotheses or theories.
Explaining Phenomena
Finally, once general laws and theories are established by testing them against specific instances of phenomena, the explaining of other specific instances consists of determining or hypothesizing the appropriate specific conditions. This is true in physics and chemistry and is especially true of the applied sciences. And the historical sciences may work exclusively in this mode. They include hypotheses about unique, past events which to be testable, must have observable consequences for the present. Here the emphasis in the testing procedure is shifted from the general laws to the specific conditions; the general laws and theories are taken for granted and the specific conditions are tested.
Repeatability
What about repeatability in the historical sciences? It is the observations that must be repeatable, if only to establish their validity independently of any one person's authority. This does not mean that the hypothetical mechanism or the phenomena concerned must be repeatable or reproducible. In the experimental lab where the phenomena being studied are short-lived and transient, it is usually necessary to reproduce them in order to repeat the observations. But scientists must wait for the reoccurrence of natural phenomena, such as eclipses, earthquakes, seasonal biological phenomena, etc. Yet if a phenomenon is a stable, more or less permanent long-time condition, observations may be repeated anytime. A geologist may return to a geological formation to repeat or make new observations. An anatomist or paleontologist may reexamine a museum specimen, either corroborating or refuting someone else's previous observations.
An Evolutionary Example
In figure 4 I have listed a number of the general consequences of the Darwinian mechanism and they constitute a fairly rigorous set of requirements which present ample opportunities for falsification. Yet in every case where there is sufficient knowledge of the specific conditions—in the form of fossil evidence—the facts are in correspondence with these general predicted features. An example:
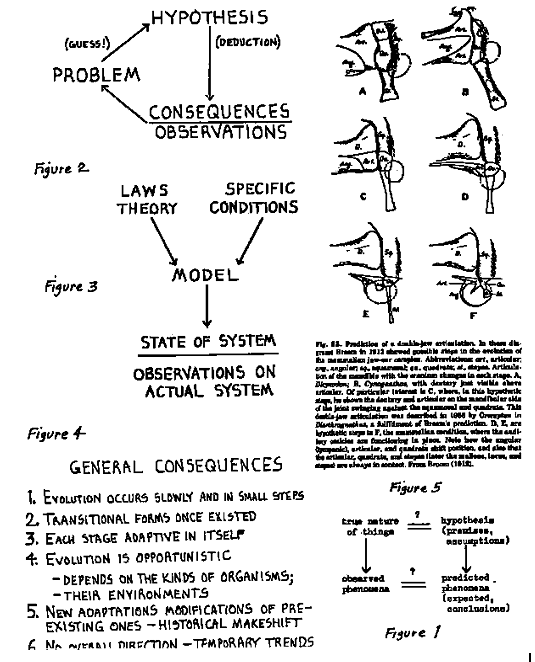
In the late 19th century, on the basis of evidence from comparative anatomy and embryology, it was hypothesized that mammals evolved from reptiles. This was independently tested (and confirmed) by the discovery of the mammal-like reptiles. Because of the relative abundance of the fossil evidence, the South African paleontologist Broom, on the assumption of natural selection, in 1912 predicted the nature of the necessary transition forms in the evolution of the mammalian jaw joint from the reptilian condition (figure 5). This included a phase where a double jaw joint, the reptilian and the mammalian ones together, occurred. It was almost 50 years later before the predicted fossils represented by Diarthrognathus and Morganucodon were actually found! This forecast of the kind of fossil that should be found is an example of the observable logical consequences possible in evolutionary theory and characteristic of science.
Hard And Soft Sciences
The oft-made distinction between the "hard" sciences (physics and chemistry) versus the "soft" sciences (biology, etc.) is based on the fact that the hard sciences have produced "universal" laws and theories admitting of no exceptions in contrast to the generalizations of the soft sciences which many authorities are even reluctant to call laws. This is often explained as a result of the hard sciences being older and more "mature" and possibly some day, ecology, for example, may produce universal laws of its own. The difference between hard and soft science, however, stems from the nature of the systems the various branches of science deal with. Physics (especially, and chemistry to some extent) restrict themselves to the simplest unvarying universal systems. Hydrogen atoms are universal and virtually identical, the variations being limited and reversible. Contrast this to a sexually-reproducing species, where virtually every individual, past and present, is unique in a great many ways; or an ecosystem, no two of which are the same. Thus, because of the nature of the subject matter, the life sciences, etc. will always produce generalizations of limited applicability and many exceptions. (Actually whenever a physicist studies a complex physical system it is separated from physics into a science of its own, such as geology and all its branches, meteorology, fluid dynamics, astrophysics, planetology, etc., all of which have features of "soft" science.)
The Mathematical Nature Of Fundamental Science
The pioneers of science—Copernicus, Kepler and Galileo—were all neoplatonists or neopythagoreans and these influences guided them in proposing mathematical hypotheses. The tremendous success of these initial researches—culminating in Newton—set off the activity of scientists and philosophers in search of mathematical laws and explanations involving the motions of particles and the properties of fields that continues to the present day.
Chance, Law And Randomness In Science
The interpretation of a chance event as one occurring in the absence of any antecedent causes is used only in some aspects of quantum physics. Otherwise a chance event or factor is one having a multiple of causes or initial conditions, most or all of which are unknown, usually for practical reasons. A large series of such events may exhibit a pattern and a probability of occurrence described by statistical laws (binomial, normal distributions, etc.)
Random has two meanings: (1) alternate outcomes of a type of event have equal probabilities of occurrence. Thus a random sample is one chosen in such a way that every individual in the population had an equal chance of being included in the sample. (2) Different events are independent, i.e. the causal chains leading up to several events have no links in common. Thus the occurrence of mutations are random with respect to the adaptiveness of the resulting phenotypes. A deterministic model explains a phenomenon or describes a system completely in terms of definitely known causes. A stochastic model includes chance factors obeying certain stated statistical laws.
There is usually a degree of inaccuracy or uncertainty and irreproducabilty in observations. Counts (ex: the number of eggs in a bird's nest) may be accurate but measurements (ex: the weight of a mouse) are usually approximate to some degree because of the limitations of the measurement device and/or procedure. Repeated observations of the same phenomenon may also display variation because each observation is the result of many factors, some of which may be beyond the control of the investigator and some of which may even be unknown. This is especially true with the complex systems of the "soft" sciences such as biology where each individual organism is unique in many ways. Thus there always appears to be a chance component to such observations.
The Role Of Statistics
When quantitative phenomena are being investigated, statistical methods allow the investigator to analyse observational data taking into consideration the chance components associated with the data. It is usually assumed that the chance elements are random in some fundamental way. The variation in observations may be summarized by descriptive statistics, deductions may involve mathematics and probability theory while the testing (falsification) procedure will involve experimental design and statistical hypothesis testing.
|
References
Aulie, R. P. 1975. The Origin of the Idea of the Mammal-like Reptile. III. The Mammal-like Reptiles. Amer. Biol. Teacher. 37(1): 21-32. (Jan.), (See esp. pp. 25-27 for jaw transition hypothesis.)
Baker, J. J. W. and G. E. Allen. 1968. Hypothesis, Prediction and Implication in Biology. Addison-Wesley.
Bronowski, J. 1960. The Common Sense of Science. Pelican Books A507. 1973. The Ascent of Man. Little, Brown and Co.
Burtt, E. A. 1932. The Metaphysical Foundations of Modern Science. Doubleday Anchor Book A41.
Campbell, N. 1952. What is Science? Dover Pub. Co. (Distinction between law and theory.)
Crombie, A. C. 1959. Medieval and Early Modern Science (2 vols.) Doubleday Anchor Books A167a, b.
Gardner, M. 1969. Mathematical Games (Simplicity as a scientific concept: Does Nature keep her accounts on a thumbnail?) Sci. Amer. 221(2): 118-121 (Aug.) 1976. Mathematical Games (On the fabric of inductive logic, and some probability paradoxes) Sci. Amer. 234:(3): 119-122 (Mar.)
Kitcher, P. 1982. Abusing Science. M.I.T. Press. (See esp. pp. 50-53, 69 for "Darwinian histories.")
Naylor, B. G. and P. Handford. 1985. In Defense of Darwin's Theory. Bioscience 35(8): 478-484. (Sept.)
Popper, K. R. 1957. The Poverty of Historicism. Boston. The Beacon Press (See esp. pp. 106, 143-147 for the historical sciences, pp. 122-124 for specific conditions, pp. 131-134 for testing and prediction.) 1959. The Logic of Scientific Discovery. Basic Books, Inc. N. Y. 1963. Conjectures and Refutations: the Growth of Scientific Knowledge. London. Routledge and Kegan Paul. (See esp. pp. 339 fol. for conditional predictions.) 1963a. Science, Problems, Aims, Responsibilities. Federation Proceedings. 224: 961-972 (July/Aug.), (An authoritative summary of Popper's views.) 1972. Objective Knowledge. An Evolutionary Approach. Oxford. Clarendon Press. (See esp. pp. 44 for truth, 355 for independence of testing.)
Ruse, M. 1979. The Darwinian Revolution. Univ. of Chicago Press. (See esp. pp. 58-59, 179-180, 279 bottom, for consilience.)
Salmon, W. C. 1973. Confirmation. Sci. Amer. 228(5): 75-83 (May).
Sokal, R. R. and F. J. Rohlf. 1981. Biometry. 2nd Ed. Freeman. San Francisco.
Stent, G. S. 1972. Prematurity and uniqueness in scientific discovery. Sci. Amer. 227(6): 84-93. (Dec.)
Article on Newton, Encyclopaedia Brittanica. 1946. 16: 362C.
Galilean Satellite; In Science and the Citizen. Scientific American 243(3): 90-92. (Sept. 1980); Did Galileo see Neptune? Science News 118(15): 231. (Oct. 11, 1980); What's bothering Neptune? Science News 119(5): 68. (Jan. 31, 1981).
The above account of Science is based in part on a paper "The Evolution/Creation Controversy and the Nature of Science" presented by F. J. Sonleitner at an Iowa Academy of Science Symposium, April 23, 1983, at Luther College, Decorah, Iowa.
The Fish-Amphibian Transition
The skeletal organization of certain crossopterygian fishes and that of early amphibians was extremely similar, the main difference being in the limbs. Ichthyostega is an intermediate form in every way—skull, axial skeleton, pectoral and pelvic girdles, the flipper-like hind leg. It also had fish scales, a fishlike caudal fin on its tail, a lateral line system, and an operculum or gill cover, presumably covering sets of gills. This and several related fossils are described by the paleontologist Jarvik as "four-footed" fishes.

|
If one objects that Ichthyostega already had "feet," it should be pointed out that the lobe-fin of the crossopterygian fish is already half-fin, half-foot. It has all the structures of the leg itself. It only needs reorganization of the peripheral elements into toes and a reorientation or rotation of the limb so the "feet" point forward.
The modern day Latimeria swims with a motion of the paired fins that resembles a trotting gait, that is, the front right fin works in tandem with the left rear and vica versa. The front fins are quite flexible and can be rotated 180 degrees.
Diagrams are from:
Schmalhausen, I. I. 1968. The Origin of Terrestrial Vertebrates. Academic Press. pp. 36, 55, 59 and 61.
See also: Jarvik, E. 1955. The Oldest Tetrapods and Their Forerunners. Scientific Monthly 80(3): 141-154.
Fricke, H. 1988. Coelacanths: The Fish that Time Forgot. National Geographic, 173:824-838. (June) for the swimming of Latimeria.
The Reptile-Bird Transition
 |
Features Of Archaeopteryx
- Dinosaur-like skull.
- Mandibles with typical reptilian teeth.
- Axial skeleton with many separate vertebrae, long tail, separate ribs, no sternum (possible cartilaginous), ventral ribs.
- Slender, relatively weak dinosaur-like pelvis.
- Forearms with long 3-fingered hand.
Overall, the skeleton is typical of many small- sized Pseudosuchian and Coelurosaurian reptiles (ex: Compsognathus) and quite different from that of the pigeon whose body skeleton show extensive fusion and reinforcement of parts forming a strong boxlike "fuselage" with a great keeled sternum, a lightweight skull and toothless beak.
The only reason Archaeopteryx was not classified as a dinosaur is because it had feathers. It was probably a climber, glider or weak flier.
Diagrams from:
Heilmann, The Origin of Birds, pp. 33, 166.
The Reptile-mammal Transition
The reptile-mammal transition is the most thoroughly documented vertebrate transition series and includes hundreds of species of the Reptilian Subclass Synapsida (the mammal-like reptiles)! What do the creationists say about it?
From Amphibians To Reptiles To Mammals
The fossil record throws very little light on the hypothetical evolution of amphibians into reptiles, or that of reptiles into mammals. All of them are four-legged vertebrates with similar skeletal structures and thus their fossilized remains provide little basis for distinguishing between them. Among animals living today, there are certain reptiles whose bony parts closely resemble those of certain amphibians and others that closely resemble certain mammals. The external characters and appearance, as well as the physiological functions, of amphibians, reptiles and mammals, are all vastly different from each other, but these differences need not show up in the fossil record.
The fact that it may be difficult to tell, for example, whether a certain fossil was a reptile or a mammal does not mean at all that it was transitional between the two in an evolutionary sense. If we could see the whole animal, and not just its skeleton, it would quickly be apparent which it was.
Of much more significance is the fact that each of the various orders of amphibians, reptiles and mammals appears suddenly in the fossil record, without incipient forms leading up to it and without transitional forms between it and any other order."
Morris, H. M. 1974. Scientific Creationism, p. 83-84.
Again Morris manages to get almost everything wrong! The fossil record throws much light on the actual transitions between these three classes, especially the reptile-mammal transition. Although they are all four-legged animals, there is no difficulty in distinguishing the fossil remains of the three classes, except for certain forms: the Semouriomorphs have a mixture of amphibian and reptilian skeletal characters and were first classified as reptiles, but more recently as amphibians when related taxa were found to have larval stages with external gills; and the cynodonts, many of which are so like the earliest forms classified as mammals that their classification as reptiles or mammals is arbitrary. Among living forms amphibian, reptile and mammal skeletons are all distincty different from one another.
Morris discounts the transitional forms by saying that if we could see the whole animal, we could quickly tell what it was. Now in the evolution of mammals from reptiles, not all the mammalian characters evolved simultaneously. Some were acquired earlier than others. For instance all mammals have hair, nurse their young and can regulate their body temperature by physiological means, thought not all can do the latter equally well. But the monotremes still have a totally reptilian mode of reproduction, and that of the marsupials is intermediate between the reptilian and the placental condition—their embryos have a vestigial egg-tooth and for the first two-thirds of pregnancy are encased in a thin shell and shell membranes. On the other hand, there is evidence that the cynodont reptiles had hair and were endothermic (warm-blooded). The Monotremes (the living platypus and echidna) have such a combination of mammalian and reptilian characters that at least one authority states that it makes more sense to classify them as living mammal-like reptiles. (Another says the advanced synapsids (therapsids) should be classified as reptile-like mammals.) Thus it seems likely that if we could see the whole animal, we might be more likely to mistake a furry cynodont for a mammal!
"The two most easily distinguishable osteological differences between reptiles and mammals, however, have never been bridged by transitional series. . .In some fossil reptiles the number and size of the bones of the lower jaw are reduced compared to living reptiles. Every reptile, living or fossil, however, has at least four bones in the lower jaw and only one auditory ossicle, the stapes.
There are no transitional forms showing, for instance, three or two jaw bones, or two ear bones. No one has explained yet, for that matter, how the transitional form would have managed to chew while his jaw was being unhinged and rearticulated, or how he would hear while dragging two of his jaw bones up into his ear."
Gish, D. T. 1972. Evolution The Fossils Say NO!. Creation-Life Pub. p. 58.
Reptiles have 7 bones in lower jaw. In mammal-like reptiles progressive reduction occurred in all of these but the dentary and articular. In the latest cynodonts, The dentary is the functional lower jaw, but the articular is still present as the jaw joint. The angular is present with a crescent shaped projection supporting the tympanum. The remaining 5 bones are reduced to tiny slivers forming a rod in a groove on the inner side of the dentary. The fossil Morganucodon, classified as a mammal, has a similar jaw except that the dentary has also established contact with the squamosal bone, so that the typical mammalian jaw joint is also present. Morganucodon and several related genera in the mammalian order Docodonta retain the reptilian lower jaw and form the new subclass Eotheria. We can't say for sure how many other of the early Mesozoic mammals had reptilian-like lower jaws because most of them are known only from teeth. Present day mammals have only one bone in the lower jaw, the dentary, but the articular is present as the malleus, the angular as the tympanic and the prearticular as the anterior process of the malleus. Thus Gish's imagined transitional series was not how it happened. Three of the original seven jaw bones disappeared; the three associated with the ear, along with the quadrate, moved to the head as a unit, leaving the dentary. The jaw never had to be unhinged and rearticulated. Broom described and predicted the actual jaw-joint transition forms in 1912 and Watson described them at length in a 1951 book. That Gish is apparently unaware of this literature documents his ignorance, not any unsolved problem of mammalian origins.
The earliest synapsids (Pelycosaurs) from the Upper Carboniferous and Permian were reptile-like. Later forms acquired incisor and canine teeth for stabbing and cutting up prey. The later synapsids (therapsids) in the Triassic acquired premolar and molar teeth, expansion of the dentary bone eventually forming a dentary-squamosal (mammalian) jaw joint in the latest forms, mammal-like skull and jaw muscle arrangements and secondary palate for chewing. Concomitant with these was a change from the reptilian alternate tooth replacement to diphylodonty (single replacement of milk teeth by permanent incisor, canines and premolars with molars added consecutively from front to back.) The post-cranial (body) skeleton also illustrates acquisition of mammalian traits associated with more efficient locomotion. All these changes first appear as minor modifications of reptilian anatomy and gradually over time became better expressed and more pronounced.
On the basis of various lines of evidence (geographic location in the higher latitudes of the Permian and Triasic world, their high predator/prey community ratios, their bone histology, indications on the skulls of pits for whiskers (and hence hair), grooves and depressions for sweat glands moistening the nose, presence of mammalian ethmoid turbinal bones, the absence of ribs in the lumbar region possibly indicating the presence of a diaphram) it is suggested that therapsids were endothermic and hairy. The more advanced ones may have nursed their young.
Hearing is often associated with the lower jaw in reptiles. In the modern lizard, part of the tympanic membrane is attached to a backward projection of the lower jaw, so that chewing affects hearing. Snakes have no tympanum yet they can hear by vibrations picked up and conducted by the jawbones. The earliest synapsids (which were not very mammal-like) also lacked of a tympanum. This was lost due to a reorganization of the back of the skull which occurred in the stem reptile (Cotylosaur) ancestors of the synapsids and obliterated the otic notch. In later synapsids, the angular bone developed a crescentric posterior margin which apparently bore a tympanum on the side of the lower jaw. This tympanum was also in contact with the articular and sounds were transmitted via the articular, quadrate and stapes to the inner ear. When the mammalian jaw-joint was formed next to the reptilian one, those bones associated with hearing became very small and migrated (along with the tympanum) off the jaw onto the head.
If you think that it is ridiculous that a reptile might have an eardrum on its lower jaw and that the jaw joint was involved in hearing, consider that living amphisbaenid lizards hear with a flap of skin on the lower jaw which communicates with the inner ear by a long cartilaginous extension of the stapes that crosses the jaw joint!
If you doubt the evidence of comparative anatomy involving the mammalian auditory ossicles, their associated muscles and innervation and blood vessels that identifies them with reptilian jaw elements, consider their embryological development. In all mammals they first form in conjunction with the lower jaw and only later move into the ear region. The newborn marsupial, which is hardly more than an embryo but which needs functional jaws to fasten to the nipple in the pouch, has a functional reptilian jaw joint (quadrate and articular)! Only after the dentary bone grows and contacts the squamosal bone of the skull forming the mammalian jaw joint, do the quadrate and articular move into the ear and take up the functions of incus and malleus, respectively. Prior to this functional shift, the embryonic jaw bones closely resemble those of a mammal-like reptile.
What the creationists must explain is: (1) why the Creator made a complete series of "transitional" forms between the reptiles and mammals and (2) how the great cataclysmic, chaotic, destructive Flood managed to sort out and deposit these forms in a perfect depositional time sequence in such beds as the Karroo formation in South Africa. Isn't it paradoxical that the creationists insist that order cannot come from chaos, yet that's exactly what they expect us to believe when they claim that the Flood produced such varied and often finely-structured sediments containing "evolutionary" sequences of all forms of organisms? Finally, to explain the embryological evidence, we might require them to explain why mammals and man are produced by developmental systems derived and modified from reptilian ones? If man is made in the Creator's image surely he ought to have a unique developmental system. Possibly that of the mammals and reptiles might be derived from man's by a lazy creator lacking in originality, and by a stretch(!) of the imagination, we might conceive that the mammalian earbones might move into the reptile jaw to become jaw elements, but the reverse?
References:
Aulie, R. P. 1974. The origin of the idea of the mammal-like reptile. American Biology Teacher. 36(11): 476; 36(12);545; 1975. 37(1): 21. (A good general account; describes Broom's predications about the jaw joint transition.)
Bakker, R. T. 1975. Dinosaur renaissance. Scientific American 232(4): 58-78 (April). (Evidence for endothermy in Triassic mammal-like reptiles.)
Brink, A. A. 1956. Speculations on some advanced mammalian characteristics in the higher mammal-like reptiles. Palaeontologia Africana, IV: 77-95. (Evidence for hair, sweat glands, single replacement dentition, possibly nursing.)
Cain, J. A. 1988. Creation and mammal origins. Journal of Geological Education 36(2): 94-105. (A good general account; answers Gish's "creative quotations" and other inaccurate comments about the subject.)
Carter, G. S. 1967. Structure and habit in vertebrate evolution. Univ. of Washington Press. (See chapter XVII for the characters of the Monotremata.)
Crompton, A. W. and Pamela Parker. 1978. Evolution of the mammalian masticatory apparatus. American Scientist 66(2): 192-201
Gans, C. and E. G. Wever. 1972. The Ear and hearing in Amphisbaenia (Reptilia). Journal of Experimental Zoology 179: 17-34.
Hopson, J. A. 1987. The mammal-like reptiles: A study of transitional fossils. American Biology Teacher 49(1): 16-27. (General review with illustrations and critique of Gish.)
Hotton, N., P. D. MacLean, J. J. Roth and E. C. Roth (Editors) 1986. The ecology and biology of mammal-like reptiles. Smithsonian Inst. Press. Washington and London. x + 326 pp. (21 papers on a variety of subjects.)
Kermack, D. M. and K. A. Kermack. 1984. The evolution of mammalian characters. London, Croom Helm. (General review with discussion of teeth, chewing, hearing and Mesozoic mammals.)
Kermack, K. A., F. Mussett and H. W. Rigney. 1973. The lower jaw of Morganucodon. Zoological Journal of the Linnean Society 53: 87-175 and 1981. The skull of Morganucodon. Ibid 71: 1-158.
Lillegraven, J. A., Z. Kielan-Jaworoska and W. A. Clemens (Editors). 1979. Mesozoic Mammals. The first two-thirds of mammalian history. Univ. of California Press. (See esp. chapter 3 on mammal origin and chapter 13 on reproduction.)
Macintyre, G. T. 1972 The trisulcate petrosal pattern of mammals. Evolutionary Biology 6: 275-303. (See p. 278 for suggestion that Monotremes are mammal-like reptiles.)
McLoughlin, J. C. 1980. Synapsida: A New Look into the Origin of Mammals. Viking Press. (See esp. the chapter: The Making of the Mammals)
Olson, E. C. 1958. The evolution of mammalian characters. Evolution 13: 344-353.
Watson, D. M. S. 1951. Paleontology and modern biology. Yale Univ. Press. (See esp. chapters 5 and 6. See p. 139 fol. for the lizard ear. A good book for paleontological methodology and examples of predictions about fossils on the basis of evolution and the observed fossil trends.)
 |
Embryos And Evolution
In 1860, Darwin wrote to Asa Gray, "Embryology is to me by far the strongest single class of facts in favor of change of forms." (Gould, 1977, p. 70). The creationists apparently agree; they studiously avoid discussing this class of facts in any detail. What Darwin was referring to was a mountain of facts emmassed by the early comparative embryologists showing that early stages in organismal development are much more similar than the adult forms would indicate. These facts were generalized in the form of two major interpretations: von Baer's laws which described the development of various forms diverging from a common general starting point and recapitulation, in which early embryonic stages of "higher" forms represent stages of forms "lower" on the ladder of life or the chain of being. Both had originated in a creationist world, but could be given evolutionary interpretations. Darwin (1958, chapter 14; 1982, chapter 13) briefly discussed these views in addition to proposing an explanation of his own. Later the German zoologist Ernst Haeckel advanced an evolutionary interpretation of recapitulation. Because present day creationists so quickly point out that recapitulation has been disproved, it is interesting to point out that the notable 19th century geologist, Louis Agassiz, embraced and expanded a creationist doctrine of recapitulation!
Why should early stages of vertebrate embryos of all classes resemble each other? Certainly all start from a single-celled zygote and go through cleavage (division of the zygote into the many small cells forming the blaustua) and gastrulation (formation of a three-layered embryo from the single layered blastula). But the parallels even extend beyond gastrulation, neurulation (neural tube formation) and into the organogenesis stage. Thus even terrestrial vertebrate embryos develop notochords, pharyngeal pouches, clefts and arches; their hearts and circulatory systems are very fish-like and undergo radical transformations, mammals display the same highly modified gastrulation process occurring in the large-yolked reptile and bird eggs; they even develop large empty yolk sacs; human embryos go through a tailed stage, etc.
The Developmental System
To understand why this must be so, it must be realized that what is inherited is information to produce a functioning developmental system that will transform a zygote into the fully developed organism. Evolution proceeds by making changes in the developmental system so that a modified organism is produced. Indeed the system accommodates change. It must. In sexual reproduction the offspring receives a mixture of information from both parents. The new genome is unique. Try this with automobile blueprints or parts of two versions of a computer program! Because of the sexual shuffling of instructions, the "manufacture" of an organism is fundamentally different from the manufacture of, say, an automobile. In our factories, parts are made in isolation. They first come together on an assembly line and fit because they were all made according to rigid specifications. The parts of an organism develop in situ, not in isolation, and constantly interact in a variety of ways to insure the production of a functional individual in spite of the mixing of "specifications" from unlike parents. Sex was invented early; virtually all eukaryote organisms have it. Thus all multicellular organisms possess developmental systems that are flexible to some extent.
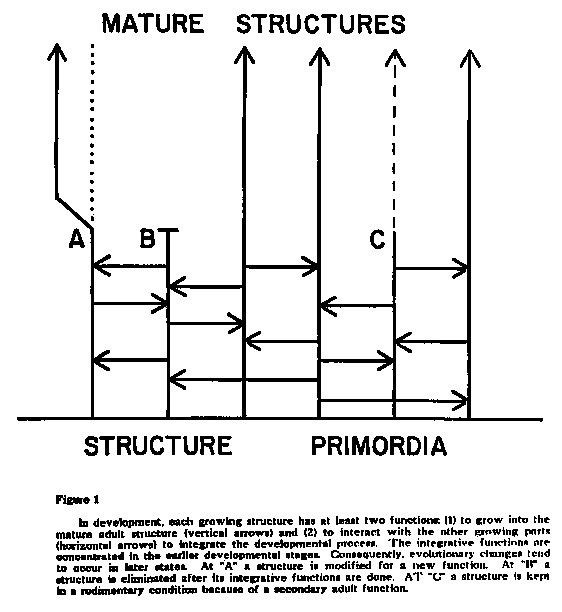
The experimental investigation of developmental systems is technologically demanding; they are very complex and very tiny (the human zygote is about the size of a pinhead) and much of their operating mechanisms are at the molecular level. Thus even with the modern techniques of molecular biology, progress in understanding them is painfully slow. Although we have an idea of the basic principles involved, there are many details still to be discovered. The principles relevant to understanding the link between development and evolution are presented in a highly simplified fashion in Figure 1. All developing structures and organs have at least two functions: (1) the primary function of becoming the operating part of the adult and (2) the secondary function of interacting with the rest of the system for the purpose of integrating it as the organism develops.
The simplest multicellular organisms, such as certain sponges and the freshwater hydra, can be teased apart into their individual cells and, left to themselves, the cells reassociate themselves into an organism (Balinsky, 1970, p. 667) In some cases mixtures of the cells from two related species can even recognize their own species. This same property is retained by the cells of various embryonic organs in complex forms (ex: kidney cells of mice; Moscona, 1961). Much of the organization in more complex forms is controlled by the process of induction. One part initiates the development of adjacent parts. Thus the presence of the notochord, which underlies the developing nervous system, is necessary for the successful differentiation of brain from spinal cord. The eye cup, growing out from the brain, induces the skin epithelium above it to form a lens; the latter in turn induces the formation of the cornea; the retinal tissue of the eye cup induces the lens material closest to it to form the lens fibers. Sometimes there is reciprocal induction. The mammalian kidney derives from two different rudimentary tissues. When these tissues grow into contact, the nephrogenic cord induces the ureteric bud to form the collecting tubes and the latter induces the former to develop the secretory tubules (Sussman, 1960, p. 80 fol.; Karp and Berrill, 1981, p. 421; Balinsky, 1970, p. 453).
Thus 20th century experimental embryology has shown that, after the main morphogenetic movements have occurred and organ and structure differentiation sets in, the system is a complex network of interacting parts. In the more advanced stages the interactions become more local and eventually the embryonic mechanisms are replaced by the nervous and endocrine systems which take over the task of keeping the organism integrated although some direct interactions persist into adult life, such as the interaction of bones, muscles and tendons; their precise form is strongly dependent upon the tensions and pressures they experience through their interactions (Weiss, 1939, pp. 448-457; Haldane and Huxley, 1927, pp. 184-187).
Clearly it would be difficult to make successful random changes in earlier stages than in the later ones. Because of the web of interactions, genes activated earlier in development are more pleiotropic (having more multiple effects) than those activated later and hence more likely to do something detrimental. Thus most successful changes come at later stages. For similar reasons, "new" organs and structures arise as modifications of old ones and not as completely new additions. (A in Figure 1.) Thus wings evolved from modified forelimbs and not as projections from the back as in angels, fairies and the mythical Pegasus. The secondary, integrative functions are very hard to modify or replace wihout instantaneous extensive reorganization, thus modifications occur later in development or if a structure or organ is eliminated, like teeth in birds, turtles, anteaters, baleen whales, etc. the early stages will be retained in some sort of rudimentary or vestigial state (B in Figure 1). Others (C in Figure 1), whose anatomy still reflects a primary function now eliminated, may persist into the adult stage because of secondary adult functions.
Examples Of The Embryological Evidence
One of the most amazing and interesting embryological phenomena in the vertebrate embryo concerns the pharyngeal gill clefts that characterize all chordates (Balinksy 1970, p. 521 fol.; Karp and Berrill, 1981, p. 421 fol.). In the invertebrate chordate (Tunicates and Amphioxus) they are part of a filter feeding apparatus. In the aquatic vertebrates they bear respiratory gills. They are formed by pharyngeal pouches growing outward that induce the formation of corresponding branchial grooves or clefts that grow inward to meet them. These two sets of structures cause the neural crest cells, migrating downward from the sides of the nerve cord to be channeled between them and form the gill bars and associated arch tissues. Several of the aortic arches so formed are incorporated into the circulatory system of terrestrial vertebrates; other tissues of the third and fourth pouches form the parathyroid and thymus glands. The hyoid skeleton and various cartilages of the larynx, trachea and thyroid are derived from arch material. Why would a Creator have these diverse structures all develop from a set of embryonic structures that develop further to form the pharyngeal gill slits in fishes and amphibian larvae? (Temporary slits do form in reptile and bird embryos and occasionally in mammals—that is, the thin membrane between pouch and groove becomes perforated but soon after closes again.) The evolutionary answer is that these organisms have necessary structures that evolved from precursors in their fish ancestors that were part of these pharyngeal structures; hence their entire elimination would be fatal to the organism.
Another bizarre example involves the growth of the main venous system. The lowest vertebrates have two main veins, the posterior cardinal veins extending backward from the heart. These appear in the embryo and grow to their adult condition in a direct manner. Mammals have a single such vein, the vena cava, which develops in anything but a straight-forward fashion. Reflecting the many evolutionary changes that occurred in the development of this venous system, including the formation of a renal portal system in fishes, its abandonment in tetrapods and the formation of adult kidneys from more posterior tubules of the basic nephric system in mammals, the venous system of the mammal starts out as paired cardinal veins, and only after a series of complicated replacements, the single vena cava develops as an amazing patchwork (Balinsky, 1970, p. 478 fol.; Arey, 1946, p. 350 fol.). If it were made by plumbers, it would be a bewildering complex of joints and branches. Surely an intelligent Creator could have planned better!
Some structures may have multiple primary functions. The yolk sac in reptiles, birds and mammals is an example. Vertebrate embryos produce ventral extensions of the developing gut to envelop the yolk so that it may be utilized for nourishment while the embryo is developing in the egg. Eggs with large amounts of yolk allow for longer incubation times and the organism hatches in a more advanced state of development. Those with small amounts of yolk hatch early either as free-living larvae that can feed themselves (ex: frog tadpoles) or are fed (by placental connections with the mother as in most mammals; from nipples in the mother's pouch in marsupials). Reptiles and birds produce large, yolky eggs and the embryos have large yolk sacs that grow and become vascularized precociously. One of the earliest sources of blood cells is in the developing yolk sac membrane. Mammals have virtually no yolk in their eggs, yet the embryo quickly produces a large empty yolk sac, clearly not to enclose yolk but to provide an early source of blood cells and in many forms to form a temporary early connection with the mother as a choriovitelline placenta until the slower growing allantois (which stores uric acid wastes in bird and reptile embryos) can establish the more permanent chorioallantoic placenta (Arey, 1946, p. 80 fol.).
The Hueristic Value Of The Embryological Evidence
The conservativeness of the early stages of development can shed light on homologies and phylogenetic relationships. This possibility, codified in Haeckel's biogenetic law, spurred the growth of comparative embryology by late 19th century embryologists investigating problems of phylogeny. For example, the sucking mouthparts of lice (Insecta: Order Anoplura) which comprise three sets of piercing and sucking stylets, are so different from the mouthparts of other insects that the homologies between the structures is not apparent. Yet when one examines the early development of a louse, it becomes apparent that the dorsal stylets form from the maxillary lobe, the ventral stylets form from the labial lobe and the intermediate stylets are the hypopharynx. The early development of certain arthropods show that the coxal glands correspond to tiny coelomic sacs and nephridial tubes of annelids; and that the book lungs of scorpions correspond to the book gills of the horseshoe crab that have withdrawn into the body. Wingless fleas have wingbuds in the pupal stage and arrangements of sclerotized plates on their thorax characteristic of winged insects. This is evidence that fleas are secondarily wingless, having evolved from winged ancestors. In contrast are the apterygote insects which are primarily wingless. They never show wingbuds and the pleural sclerites of their thorax are similar to those on the abdomen and to those of other wingless arthropods such as the Chilopoda (centipedes). The larval form of the ascidians (sea squirts) revealed that they were chordates, not molluscs, while the larval form of the crab parasite, Sacculina indicates that it is a form of barnacle. Similarly the Ammocoetes larva of the lamprey shows the relationship between the vertebrates and the cephalochordates (Amphioxus). And the pharyngeal gill structures of terrestrial vertebrates indicate their kin to the aquatic vertebrates.
Other Patterns Of Developmental Modification
The gradual divergence of development of descendant forms from that exhibited by forms still retaining ancestral features is not the only way that evolution can modify developmental systems. The timing of various processes may change. DeBeer (1940) and Gould (1977) discuss such heterochrony at length. An example of heterochrony is paedomorphosis, the retention of juvenile ancestral characters by later ontogenetic stages of descendents. This process is envoked to explain why humans resemble juvenile apes in many morphological features much more than they do adult apes (Gould, 1977). (The author Aldous Huxley was so fascinated by this idea that he turned it into a plot for a novel: After Many a Summer Dies the Swan. New York: Harper and Bros.) The reverse process, the accelerated appearance of ancestral characters into earlier ontogenetic stages of descendants, mimics recapitulation, as in the opposum embryo, where the articular and quadrate bones form a functional reptilian jawjoint so that the oppossum can attach to its mother's nipple, those two bones later transforming into the malleus and incus earbones when the mammalian dentary-squamosal jawjoint finally forms (Lillegraven et al, 1979, p. 265; Watson, 1951, p. 156).
The fully functioning wing of adult insects contains little living material, hence these adults cease growing and molting. Thus the life cycle includes flightless, juvenile growth stages and the flying adult dispersal and reproductive stage. In the holometabolous insects (those with complete metamorphosis) these two main stages have diverged greatly both in ecology and morphology and the developmental system has virtually split into two. This is most evident in the flies (Diptera) and butterflies and moths (Lepidoptera). The caterpillar contains within it little islands of undifferentiated cells, the imaginal disks. In the pupal stage, most of the larval tissues are destroyed and the adult butterfly grows from these imaginal disks. Other insect orders exhibit this double developmental system to varying lesser degrees or not at all. (Saunders, 1982, p. 401 fol.).
Creationists And Recapitulation
Creationists attempt to explain away this embryological evidence by saying that recapitulation has been disproven (Morris, 1974, p. 77; Kofahl, 1980, p. 103). The fact that a hypothetical explanation of these embryological phenomena has been found wanting does not means that the phenomena do not exist. The phenomena do exist, and the evolutionary explanation based on the findings of experimental embryology as outlined above was proposed early in this century (Needham, 1930) and is accepted by contemporary workers (Balinsky, 1970, p. 590, Waddington, 1966, pp. 5-7), and correctly described by the better biology textbooks (examples are Simpson, Pittendrigh and Tiffany, 1957, pp. 352-354 and Keeton, 1972, pp. 551-552 ). A review of eighteen of the latest biology/zoology and eight developmental biology/embryology textbooks reveals an unfortunate trend to slight or even eliminate "traditional" subjects like embryonic similarities and expand upon more "modern" topics such as molecular biology and molecular genetics. Also, many of the modern authors still refer to these embryological phenomena and their evolutionary explanation as recapitulation. Two exceptions are Hopper and Hart (1985, pp. 3-5) and Hickman, Roberts and Hickman (1988, pp. 9, 125-126, 456-457).
Creationists give the impression that recapitulation was a necessary corrolary to evolution invented by Ernst Haeckel (see Gordon, 1987, p. 7). Actually, the idea of recapitulation was held by a number of pre-evolutionary biologists starting with C. F. Kielmeyer in 1793, Lorentz Oken in 1809-1811, J. F. Meckel in 1811 and Etienne Serres in the 1820's and 30's. These authors all incorporated recapitulation into non-evolutionary philosophies, explaining it on other bases such as the unity of nature's laws or relating it to the chain of being or scale of nature. It was later championed by Louis Agassiz as illustrating a three-fold parallelism between embryonic growth, the structural gradation in adult forms and the geologic succession of fossils that represented the history of a particular type (Gould, 1977, chapter 3; Mayr, 1982, p. 469 fol.). Von Baer rejected recapitulation and instead stated that general characters of a group developed first and the more specific ones distinguishing the members of the group appeared later. In later editions of the Origin, Darwin mentions the work of von Baer, Muller (whose views were similar to Haeckel's) and Agassiz but avoids their explanations. He considers the phenomena as representing a law of embryonic similarity and offers his own explanation—that most successful variations appear later in the development of the individual. This fits in with the modern explanation given above. Darwin, of course, wasn't aware of the web of interactions found by the 20th century embryologists that explains why successful modifications tend to occur later in development. Those same embryologists falsified Haeckel's extreme brand of recapitulation—that all evolutionary changes are added to the end of the developmental sequence, causing individuals to develop through a sequence of the adult stages of their ancestors.
Creationists And The Similarities Of Early Development
Another creationist argument is to claim that the resemblances of early embryonic stages are trivial and superficial: ". . .and since furthermore many of the structures to be developed must be somewhat similar (limbs, head, etc.), it would be natural that the developing embryos would look much alike for the initial stages of their development." (Morris, 1974, p. 73; see also Morris, 1967, p. 24). Another argument is to claim to satisfactorily explain these seemingly anomalous embryonic structures by simply pointing out functions for them. Both Morris and Kofahl list the various derivatives of the pharyngeal pouches and conclude that they reflect "careful planning and design" (Morris, 1974, p. 77); ". . .the facts agree quite logically with the creation viewpoint rather than with an evolutionary explanation." (Kofahl, 1980, p. 104). Evolution also explains their continued existence on the basis of function. But evolution goes further and explains why these structures have the shapes and anatomy that they do. On the other hand, why would a Creator choose to provide blood cells to the early stages of a mammal embryo with an empty yolk sac? Why couldn't the omniscient and omnipotent Creator speed up the growth of the allantois so that a temporary yolk sac placenta is unnecessary? Why should a terrestrial vertebrate embryo produce pharyngeal pouches to induce branchial grooves and gill clefts that in turn induce gill bar tissue to serve as the source of thymus and parathyroid glands which then have to shift their location downward and backward? Evolution explains these incredibly conservative "design" features. The mechanism of mutation and natural selection couldn't proceed in any other way but make use of structures already present and part of the integrative network. Natural selection works more like a tinkerer than an omnipotent designer (Jacob, 1977). On the other hand, special creation would suggest that the egg should develop to the adult by the most direct route without the extraordinary detours that one actually finds (Mayr, 1982, p. 470).
Creationists And Vestigial Organs
Creationists explain vestigial organs by claiming that they have a function, hence they are not vestigial (Morris, 1974, p. 75; Kofahl, 1980, p. 102; Moore and Slusher, 1974, p. 435), or alternately that they are examples of decay and deterioration, not evolution (Kofahl, 1980; Morris, 1963, p. 44; Morris, 1968, p. 53; Morris, 1974, p. 75). Evolution explains the persistence of vestigial organs which no longer perform their primary function. They may either have an additional primary function, like in the yolk sac example above, and the lymphoid tissue in the appendix (Moore and Slusher, 1974, p.435) or may be retained in embryo because of their secondary integrative function (Yablokov, 1966). Why should the jaws of specially-created toothless animals need teeth (baleen whales) or tooth buds (birds) to insure proper growth of the jaws (Howe, 1985)? And why should birds have a complete set of genic instructions for making fully formed teeth (Kollar and Fisher, 1980) when it is normally never used. Evidently the operation of the tooth-forming gene complex is suppressed in birds but not eliminated because the production of tooth buds may be necessary for the proper growth of the jaws. Similarly mechanisms persist to make fully formed hind limbs as occasionally occurs in whales (Edwords, 1983), or three toed feet as sometimes occurs in horses (Gould, 1980, McGowan, 1984, pp. 147-148), or persistent cervical slits resulting in perforated necks in humans (Arey, 1946, p. 179).
Darwin proposed that vestigial or rudimentary organs resulted from disuse aided by natural selection if the organs were detrimental in the organism's new environment. Moore and Slusher (1974, p. 436) claim that evolution cannot explain vestigial organs at all: deterioration because of disuse is Lamarkian and why would "loss mutations" be selected for losing an organ that might not be beneficial but certainly was not harmful? The accepted evolutionary explanation involves the pleitropic effects of genes (see above) and the gene complexes that produce the various structures. In a cave dwelling organism, for example, selection for gene complexes producing inhanced senses of smell, touch, hearing, etc. may involve actively breaking up gene complexes for eyes because such complexes share some genes (Allee et al., 1949, pp. 672-679). The existence of such overlapping complexes may also prevent the complete elimination of a "useless" structure.
Other Creationists Arguments
In general, creationists recognize the weakness of their arguments and prefer to avoid dealing with Darwin's most compelling evidence. The Origins Two Models book (Bliss, 1976) never mentions it. The creationist biology textbook (Moore and Slusher, 1974) doesn't describe any of the embryonic similiaries in the chapter on embryology although they are cursorily mentioned in the chapters criticizing evolution (see pp. 432-436). This latter book (p. 435) also makes the inane claim that evolutionists must assume that vestigial nipples in men must have been functional in ancestral males. An organism's developmental system must produce both males and females. A vertebrate embryo goes through an early stage that has a complete set of primordia for both sexes. Once sex hormones take over, one set of structures remains vestigial while the other develops. How do creationists explain this, considering that Adam was made first and Eve was only an afterthought, made from Adam's rib? The question has long been asked: did Adam have a navel? What about nipples? or for that matter, the vestigial remains of oviducts (appendix testis) and vagina (prostatic utricle or vagina masculina)? Similarly women have vestiges of the male ducts. Like the hind limbs of whales, the degree of development of these vestiges varies a great deal from individual to individual (Arey, 1946, p. 297 fol.).
Summary
Evolutionary mechanisms tend to modify the later stages of developmental systems because of the web of interacting and integrating parts found in the earlier stages cannot easily be modified. Thus organs and structures that are modified or eliminated in the later stages are retained in the earlier stages because of their integrative functions. Some rudimentary or vestigial organs may persist into the adult stage because they have a secondary adult function in addition to the integrative one. Organs that are no longer useful may become vestigial or eliminated when natural selection breaks up the gene complexes producing these organs in order to assemble more efficient complexes for other organs that are still necessary for survival.
 |
References
Allee, W. C., A. E. Emerson, O. Park, T. Park and K. P. Schmidt. 1949. Principles of Animal Ecology. Philadelphia: W. B. Saunders Co.
Arey, L. B. 1946. Developmental Anatomy. A Textbook and Laboratory Manual of Embryology. 5th Ed. Philadelphia: W. B. Saunders Co.
Balinsky, B. I. 1970. An Introduction to Embryology. 3rd Ed. Philadelphia: W. B. Saunders Co.
Bliss, R. B. 1976. Origins: Two Models: Evolution Creation. San Diego, CA: Creation-Life Pub.
Darwin, C. R. 1958. The Origin of Species. New York: Mentor Books, New American Library. (reprint of the last edition, with an introduction by Julian Huxley).
Darwin, C. R. 1982. The Origin of Species. New York: The Penguin English Library. (reprint of the first edition, 1859, edited by J. W. Burrow).
DeBeer, G. R. 1940. Embryos and Ancestors. Oxford: Clarendon Press.
Edwords, F. , 19,, 83. "Those Amazing Animals: The Whales and Dolphins." Creation/Evolution X: 1-7.
Gordon, P. 1987. "Articles." Origins Research 10(1): 6-7. Spring/Summer.
Gould, S. J. 1977. Ontogeny and Phylogeny. Cambridge, MA: Belknap Press.
Gould, S. J. 1980. "Hen's Teeth and Horse's Toes." Natural History 89(7): 24-28. (July)
Haldane, J. B. S. and J. Huxley. 1927. Animal Biology. Oxford: Clarendon Press.
Hickman, C. P., L. S. Roberts and F. M. Hickman. 1988. Integrated Principles of Zoology. St. Louis: Times Mirror/Mosby College Publishing.
Hopper, A. F. and N. H. Hart. 1985. Foundations of Animal Development. 2nd Ed. New York: Oxford University Press.
Howe, G. 1985. "Further comments on baleen fetal teeth and functions for yolk sac." Origins Research 8(2): 13-14. Fall/Winter.
Jacob, F. 1977. "Evolution and Tinkering." Science 196(4295): 1161-1166. (10 June.)
Karp, G. and N. J. Berrill. 1981. Development. 2nd Ed. New York: McGraw-Hill.
Keeton, W. T. 1972. Biological Science. 2nd Ed. New York: W. W. Norton and Co.
Kofahl, R. E. 1980. The Handy Dandy Evolution Refuter. San Diego: Beta Books.
Kollar, E. J. and C. Fisher. 1980. "Tooth Induction in chick epithelium: Expression of Quiescent Genes for Enamel Synthesis." Science 207: 993-995. (29 February)
Lillegraven, J. A., Z. Kielan-Jaworoska and W. A. Clemens (editors). 1979. Mesozoic Mammals. The first two-thirds of mammalian history. Univ. of California Press.
Mayr, E. 1982. The Growth of Biological Thought. Diversity, Evolution and Inheritance. Cambridge, MA: Belknap Press.
McGowan, C. 1984. In the Beginning. . . Buffalo: Prometheous Books.
Moore, J. N. and H. S. Slusher. (Editors) 1974. Biology: A Search for Order in Complexity. Grand Rapids, MI: Zondervan Pub. House.
Morris, H. M. 1963. The Twilight of Evolution. The Craig Press.
Morris, H. M. 1968. The Bible and Modern Science. Chicago, IL: Moody Press.
Morris, H. M. 1967. Evolution and the Modern Christian. Philadelphia: The Presbyterian and Reformed Pub. Co.
Morris, H. M. (Editor) 1974. Scientific Creationism. San Diego, CA: Creation-Life Pub.
Moscona, A. A. 1961. "How Cells Associate." Scientific American 205(3): 142-162. (September.)
Needham, J. 1930. "The biochemical aspect of the recapitulation theory." Biological Reviews and Biological Proceedings of the Cambridge Philosophical Society V: 142-188.
Saunders Jr., J. W. 1982. Developmental Biology. New York: Macmillan.
Simpson, G. G., C. S. Pittendrigh and L. H. Tiffany. 1957. Life, An Introduction to Biology. New York: Harcourt, Brace and Co.
Sussman, M. 1960. Animal Growth and Development. Englewood Cliffs, NJ: Prentice-Hall, Inc.
Weiss, P. 1939. Principles of Development. A Text in Experimental Embryology. New York: Holt and Co.
Waddington, C. H. 1966. Principles of Development and Differentiation. New York: Macmillan.
Watson, D. M. S. 1951. Paleontology and Modern Biology. New Haven, CN: Yale Univ. Press.
Yablokov, A. V. 1966. Variability of Mammals. Moscow: Nouka Pub. (1974 translation, Amerind Pub. Co.)
(from Frank Sonleitner's critique of Of Pandas and People)